
My Sites
Log in to create or edit your sites.
IMPORTANT SERVICE ANNOUNCEMENT: Content freeze 7am June 19th until 7am June 23rd. Read more here.
Need Help? Email edtech@tufts.edu
Site-Wide Activity
-
Alissa B. Mariano is attending Seminar III. 10 years, 7 months ago
-
Brooke Traylor wrote a new post on the site Museum Studies Blog at Tufts University 10 years, 7 months ago
The Massachusetts Historical Society’s latest exhibit, “Letters and Photographs from the Battle Country: Massachusetts Women in the First World War,” tells of the Great War from a lesser-known perspective: that of […]
-
Grant Beyer is attending Seminar IV. 10 years, 7 months ago
-
Michael O. Onyekaba is attending D'16. 10 years, 7 months ago
-
Claire McCarthy is attending D'16. 10 years, 7 months ago
-
Kishan V Patel is attending D'16. 10 years, 7 months ago
-
Eric J Berger is attending D'16. 10 years, 7 months ago
-
Sabrina Coombs is attending Seminar I. 10 years, 7 months ago
-
Chiara L. Castellucci is attending Seminar III. 10 years, 7 months ago
-
Anthony Minhduc H. Phan is attending Seminar III. 10 years, 7 months ago
-
Shivam A. Patel is attending Seminar III. 10 years, 7 months ago
-
Lalita Nekkanti is attending Seminar IV. 10 years, 7 months ago
-
Sharod McKinney is attending Seminar III. 10 years, 7 months ago
-
Ann Gagliardi wrote a new post on the site Tufts SAVMA 10 years, 7 months ago
[ngg src=”galleries” ids=”23″ display=”basic_thumbnail”]

-
willie wrote a new post on the site Emotion, Brain, & Behavior Laboratory 10 years, 8 months ago
In my last post, I talked about the potential imbalance in the dopaminergic system as being part of the reason for compulsive behaviors. I loosely associated these behaviors to those found in OCD. I will continue to make this association in this post and the ones to come. This post will focus on studies that show differences in brain structure of people with and without OCD. We will see that much of the differences will correlate with regions involved in the dopaminergic system. Ultimately, my goal is to show that it is plausible that a deficiency in processing rewards, much of which takes place in the orbitofrontal cortex (OFC), is a major source of the dopamine imbalance and resulting compulsive behaviors.
Let me begin with describing some of the functionality believed to be related to the OFC. These functions include decision-making, judgements, planning, and reward processing. In this post I will focus on reward processing and goal pursuit. It is hard to fully separate these two from functions like decision-making and planning since choosing a goal is a decision-making process and then one constructs a plan for pursuing that goal. Nonetheless, I will focus here on goals and reward, and in my final post a few weeks from now I will go discuss decision-making.
Why do you pursue a goal? The goal must have some benefit, and that benefit manifests itself in the form of a reward. It the brain has difficulty in processing rewards, then it loses motivation to pursue the goal. Looking at the other side of the chicken and egg problem, if the brain is not able to focus on goal pursuit, then the benefit and reward for achieving the goal cannot be realized. It is unclear which is happening, but these are clearly linked. And naturally, the OFC is responsible for both goal pursuit and processing of rewards. Whether the problem in the OFC is related to the goal or the reward is still a mystery.
By now you may be asking if there is evidence that the OFC is different in people with and without OCD. There is actually a substantial amount of evidence using both positron emission tomography (PET) and functional magnetic resonance imaging (fMRI) showing significantly more metabolism and activation in the OFC. Harris & Hoehn-Saric (1995) using MRI images found no structural differences between controls and those with OCD. However, the PET scans showed hypermetabolism in the OFC. More specifically, PET scans have shown increased metabolic rates for the right OFC (Kwon et al., 2003).
Tests with fMRI have shown similar results. In a comparison of patients with OCD and controls, while they are in a resting state, those with OCD presented increased amplitude of low-frequency fluctuation (ALFF) in the OFC. Additionally, the ALFF values were positively correlated with total Y-BOCS scores (Hou et al., 2012). (Y-BOCS is the Yale-Brown Obsessive Compulsive Scale, a standard test used for diagnosing OCD. Try it yourself at http://psychology-tools.com/yale-brown-obsessive-compulsive-scale/)
If the OFC is partially responsible for OCD, then after treatment (pharmacological or behavioral therapy), then one would hope that the hyperactivation of the OFC would be mitigated. Nakao et al. (2005) found that after such treatment, scores on the Y-BOCS test decreased and activation in the frontal lobe also decreased.
Hopefully, I have presented a good enough case that the OFC plays a significant role in OCD. Also, as I stated earlier, the OFC is related to goal pursuit and reward processing. Are these related? (Of course! Why else would I be asking the question?) A fascinating and recent study tested patients with OCD on their ability to use goal-directed behavior. The patients and the controls learned a new task, and both were able to learn the task at similar rates. However, the OCD patients performed poorly in their ability to adapt to changes in the task and instead fell into habits (Papmeyer et al., 2011). The authors relate this to resorting to compulsive behaviors like hand-washing and checking. I interpret this as a reward anticipation has been created, but the corresponding goal cannot be identified or selected and instead the person resorts to established behaviors in hopes of acquiring the reward. Unfortunately, the reward may be nonexistent or too little, and the cycle of reward anticipation and failed acquisition persists.
There are many brain regions the have been shown to be related to OCD, including the OFC, striatum, anterior cingulate, thalamus, and caudate nucleus (Saxena & Rauch, 2000). I focus here on the role of the OFC and its functioning for reward processing and goal pursuit. In my last post I began to make a case for an imbalance in the dopaminergic pathway, and this post I begin to provide evidence that malfunctioning of the OFC may be contributing to this imbalance. The striatum is releasing plenty (or too much) dopamine but perhaps the OFC is not able to metabolize it. The reward anticipation maintains but the reward is not fully realized. This is likely related to a deficiency in adapting and pursuing goals, and instead the ritualistic behaviors are repeated.
I will continue to look into this in my next post and look at the role of memory and learning. Reward is often related to learning, so we would expect to see an impact there.
References
Harris, G. J., & Hoehn-Saric, R. (1995). Functional neuroimaging in biological psychiatry. Advances in biological psychiatry, JAI Press, Greenwich, 113-160.
Hou, J., Wu, W., Lin, Y., Wang, J., Zhou, D., Guo, J., … & Li, H. (2012). Localization of cerebral functional deficits in patients with obsessive–compulsive disorder: A resting-state fMRI study. Journal of affective disorders,138(3), 313-321.
Kwon, J. S., Kim, J.-J., Lee, D. W., Lee, J. S., Lee, D. S., Kim, M.-S., … Lee, M. C. (2003). Neural correlates of clinical symptoms and cognitive dysfunctions in obsessive-compulsive disorder. Psychiatry Research, 122(1), 37–47.
Nakao, Tomohiro, et al. “Brain activation of patients with obsessive-compulsive disorder during neuropsychological and symptom provocation tasks before and after symptom improvement: a functional magnetic resonance imaging study.”Biological psychiatry 57.8 (2005): 901-910.
Saxena, S., & Rauch, S. L. (2000). Functional Neuroimaging and the Neuroanatomy of Obsessive-Compulsive Disorder. Psychiatric Clinics of North America, 23(3), 563–586.
Papmeyer, M., Morein-zamir, S., Ph, D., Sahakian, B. J., Ph, D., Fineberg, N. A., … Ph, D. (2011). Disruption in the Balance Between Goal-Directed Behavior and Habit Learning in Obsessive-Compulsive Disorder. American Journal of Psychiatry, 168(July), 718–726.
-
Kate Dahlgren wrote a new post on the site Emotion, Brain, & Behavior Laboratory 10 years, 8 months ago
 In a recent episode of The Simpsons, “MoneyBart,” when floundering father, Homer, is confronted with his responsibilities, he decides to shirk them in order to get drunk (Long & Kruse, 2010). He justifies his […]
In a recent episode of The Simpsons, “MoneyBart,” when floundering father, Homer, is confronted with his responsibilities, he decides to shirk them in order to get drunk (Long & Kruse, 2010). He justifies his […] -
Jennifer M. Perry wrote a new post on the site Emotion, Brain, & Behavior Laboratory 10 years, 8 months ago
In February 2012, 17 year-old Thomas “T.J.” Lane entered his high school with a .22 caliber handgun, killed three of his classmates, and injured several others. Sadly, as school shootings have devastatingly become more commonplace each year in the United States, this is not the most shocking part of the story. At his sentencing in 2013, T.J. Lane not only demonstrated absolutely zero remorse, but actually seemed to experience pleasure at listening to the victim statements read by the families. Before their statements, he took off his dress shirt, revealing a t-shirt with letters written in marker, spelling “KILLER.” He continuously smiled and smirked as they described their anguish. He even went so far to tell the families about how he fantasized about their sons’ deaths (though in much cruder language than I just communicated—the video is available on YouTube, but if you choose to watch, be forewarned about the crudeness of his statements).

T.J. Lane at his sentencing
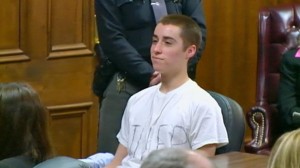
Smirking while listening to the family’s victim impact statements
There is an innumerable amount of books, television shows, movies, documentaries, etc about criminals, especially on those who inflict exorbitant pain on others. We clearly are fascinated with the topic. Why? It seems truly unbelievable that a human being could contemplate, let alone carry out, these types of actions. How could another person possess such a different moral compass than our own? How could a human being possibly be motivated to kill, sometimes without motive, and experience absolutely zero remorse? Admittedly– as truly horrible as it is– it is also fascinating, and a subject to which psychologists and neuroscientists have dedicated a good deal of research.
In my first post on the theme of “Failures in Empathy,” I described various contexts in which regular people can be motivated to demonstrate a lack of empathy towards others. This week, however, I am shifting gears to explore one of the most striking examples within the realm of empathic failure: Psychopathy. This week’s theme represents the opposite thesis of that presented in the previous post, in that it will explore the ways in which empathic failures manifest themselves in entirely atypical individuals.
Psychopathy is defined by callous, impulsive, and antisocial thoughts and behavior. Psychopaths tend to experience no fear of punishment and a lack of emotional sensitivity and reactivity (Decety et al., 2013). In other words, psychopaths are, in part, definitionally characterized by their empathic failures. But what are the mechanisms that underlie an almost constant lack of empathy? What is the difference between most of us, who require sufficient motivation to experience a small taste of empathic failure, and psychopaths, whose mere baseline seems to be represented by empathic deficiency? What made T.J. Lane so inexplicably heartless about the deaths of his victims, whereas most of us may become distressed by merely watching his reaction, let alone by imagining his cruel acts?
In order to examine the underlying mechanisms of empathic deficiencies in psychopathy, it may be helpful to parse the concept of empathy further. Empathy consists of three components: a.) an affective component, which often entails sharing the emotion of the target individual (e.g. feeling distress at another person’s distress) b.) a cognitive component, which involves the capacity to take the perspective of another person. and c.) a regulatory component, in which one is able to monitor the origins of the feelings (self vs. other) (Lamm et al., 2007). As such, empathy generally has two overarching facets: the affective (emotional) and the cognitive. While the cognitive facet allows us to recognize what the other person is feeling (and to keep in mind that the origin of the feelings stems from observing the other person), the affective facet prompts us to ‘feel’ as the other person feels.
Research has shown that psychopaths are not deficient in all components of empathy. In fact, a number of studies have demonstrated that only the affective component is implicated in psychopathy, while the level of cognitive functioning remains relatively normal (see Blair, 2005 for a review). Simply put, psychopaths realize, recognize, and fully comprehend the distress their victims are feeling. They simply, though, do not feel it themselves.
Using this general finding, Decety et al. (2013) sought to isolate the specific empathic deficiencies of psychopathic individuals. In their study, the authors took advantage of the fact that psychopaths are, indeed, capable of perspective taking. They had psychopathic individuals engage in a process called affective perspective taking (imagining the emotional reaction of another person) while viewing pictures of other people in pain. Affective perspective taking has been shown to be a powerful method to increase empathic concern towards others (Batson et al., 1997). The experimenters, however, had the psychopathic individuals engage in the perspective taking in two different ways: 1. an ‘imagine-self’ perspective (“Imagine this situation is happening to you.”) and 2. an ‘imagine-other’ perspective (“Imagine this situation is happening to someone else.”). Results showed that when taking the imagine-self perspective, the neural activity of the psychopathic individuals was similar to that of non-psychopathic individuals— brain areas typically involved in empathic and emotional processing and reactivity were activated (the amydala, anterior insula, anterior midcinguate cortex, to name a few). In the imagine-other perspective, however, activation in these areas remained in the control participants, but was nearly completely absent in the psychopathic participants. These results are important in that they show that psychopathic individuals are not completely devoid of emotional processing. When it comes to themselves, they seem exhibit generally normal emotional functioning. Doing the exact same thing, but for others, is where they fall short.
The results of the Decety et al. (2013) study were also provocative in another important way— finding increased activation of the amydala in the psychopathic individuals. The vast majority of neuroscientific research on psychopathy has shown that psychopaths show greatly reduced amygdala functioning. This makes sense, as the amygdala is most notably known for its role in fear conditioning, a paradigm in which a previously neutral stimulus is paired with a fear-inducing stimulus to generate a fear response (see LeDoux, 1996). Psychopaths have been shown to exhibit very poor fear conditioning (are very difficult to induce with fear), and thus, decreased amygdala functioning (Blair, 2007). In fact, poor fear conditioning and the corresponding decreased amygdala response in 3 year olds have been shown to predict degree of psychopathy in the same individuals as adults (Gao et al., 2010—a study I just learned about this morning in an awesome talk by Adrian Raine here at Tufts!).
If psychopaths themselves are much less sensitive to fear than normal individuals, it would not be an outlandish conjecture that they would be extremely insensitive to others’ fear. Accordingly, Marsh and Cardinale (2012) found that psychopaths were significantly impaired in identifying statements/behaviors that could cause fear (e.g. “I could easily hurt you”; “You better watch your back.”), as well as in judging the moral acceptability of these statements/behaviors. Compared to controls, psychopaths were less able to recognize fear-inducing statements (compared other emotions of ‘anger’, ‘disgust’, ‘happiness’, and ‘sadness’), and also judged the fear statements to more acceptable. Thus, while a good deal of research has shown that psychopaths are still typically able to recognize others’ emotions, ‘fear’ (a highly empathy-eliciting emotion) may be the exception.
In this post I have presented the ways in which very atypical individuals exhibit empathic failures. In the next post, I will return to a context in which empathic failures manifest in us all: when our group memberships take over.
————–
Batson, C., Early, S., & Salvarini, G. (1997). Perspective taking: imagining how another feels versus imagining how you would feel. Personality and Social Psychological Bulletin, 23, 751–758.
Blair, R. (2005). Responding to the emotions of others: dissociating forms of empathy through the study of typical and psychiatric populations. Conscious Cognition, 14, 698–718.
Blair, R. (2007). The amygdala and ventromedial prefrontal cortex in morality and psychopaths. Trends Cognitive Science, 11, 387–392.
Decety, J., Chen, C., Harenski, C. L., & Kiehl, K. A. (2013). An fMRI study of affective perspective taking in individuals with psychopathy: imagining another in pain does not evoke empathy. Frontiers in Human Neuroscience, 7, 489.
Gao, Y., Raine, A., Venables, P. H., Dawson, M. E., & Mednick, S. A. (2010). Poor childhood fear conditioning predisposes to adult crime. American Journal of Psychiatry, 167, 56-60.
Lamm, C., Batson, C. D., & Decety, J. (2007). The neural substrate of human empathy: Effects of perspective-taking and cognitive appraisal. Journal of Cognitive Neuroscience, 19, 42–58.
LeDoux J.E. (1996) The Emotional Brain. New York: Simon & Schuster.
Marsh, A. A. & Cardinale, E. M. (2012). Psychopathy and fear: Specific impairments in judging behaviors that frighten others. Emotion, 12, 892-898.
[Photos of T.J. Lane]. Retrieved October 24, 2014, from: http://www.nydailynews.com/news/national/ohio-teen-faces-life-prison-shooting-rampage-left-high-school-students-dead-article-1.1292702 and http://www.usatoday.com/story/news/nation/2014/09/11/chardon-shooter-escaped-from-prison/15486753/
-
Benjamin Heuberger wrote a new post on the site Emotion, Brain, & Behavior Laboratory 10 years, 8 months ago
A few weeks ago, as I was complaining to a friend of mine about the tremendous amount of work I had at the time, he told me, half-jokingly, to “man up”. We are all familiar with this expression and the stereotype upon which it’s based. According to this stereotype, a “real man” isn’t supposed to be psychologically weak or emotionally sensitive. This got me thinking: Is there any empirical evidence that supports the idea that women are perhaps
more in touch with their emotions, and if so, are these differences reflected in memory differences for emotionally salient stimuli? Not only does the literature suggest differences in emotional memory between men and women, but neural activation during retrieval processes seems to be different as well, suggesting that perhaps men and women don’t necessarily rely on the same neural pathways when it comes to emotional memory.Mackiewicz et al. (2006) built off these findings and found that lateral activation differences between men and women were only observed in the ventral amygdala, while no lateralized difference was observed in the dorsal amygdala. Furthermore, Cahill et al. (2004) demonstrated that this lateralization was observed not just in the amygdala, but other brain regions as well. Among women, all significant activations associated with emotional memory were located in the left hemisphere, including the left posterior cingulate, left middle temporal gyrus, and left inferior parietal cortex. Conversely, neural activation in men was generally found in the right hemisphere in similar structures.
[Credit: http://www.mybraintest.org/%5D%5B/caption%5DAs I discussed in my last blog post, emotionally salient information is generally remembered better than similar non-emotional, neutral information. By emotionally salient—specifically in an experimental context—I refer to stimuli that evoke stronger than usual feelings in an individual as determined by self-report, and in many cases, corresponding physiological changes. Yet although both men and women may remember emotionally salient information better than neutral information, there also appear to be sex differences with regard to memory of emotional information.
Canli et al. (2002) examined memory for a series of neutral and highly negative pictures after a three-week delay in a sample of men and women. Participants also rated the pictures prior to the memory task to provide their subjective evaluation of how emotionally arousing they were (on a scale of 1, calm/dull, to 9, most arousing or intense). Interestingly, women on average rated more pictures as highly arousing than men did and had better recall for emotionally arousing pictures than men did after the delay. Because this sex difference in memory was not observed for neutral pictures, it seems the superior performance by women was due specifically to the emotionally arousing nature of the pictures. Other studies have demonstrated that in a free recall task where participants were asked to recall their strongest and most emotional positive and negative memories from the last year within an allotted timeframe, women tend to report both more positive and negative memories. (Fujita et al, 1991), (Seidlitz & Diener, 1998). This finding is in line with the stereotype of women being more “in touch” with their feelings and emotions.
Sex differences in neural activity during emotionally salient memory processes may shed some light on why women seem to show have better emotional memory. Numerous studies have found differences in hemispheric lateralization of amygdala function in emotional memory. As discussed in my last blog post, the amygdala plays a selective role in hippocampal-dependent consolidation and retrieval of emotional memory. In the study described earlier (Canli et al., 2002), researchers found that recognition accuracy for maximally arousing pictures—but not for less arousing or neutral pictures—was correlated with greater left amygdala activation in women but not men. In men, greater recognition accuracy for arousing information was correlated with greater activation of the right amygdala activation. Cahill et al. (2004) also found relatively greater activation of the right amygdala in men and left amygdala activation in women and memory for emotionally arousing information.
[caption id="attachment_317" align="alignright" width="239"]
![Hemispheric Lateralization of Amygdala Activity During Successful Recall of Emotional Memory [Credit: Canli et al. (2002)]](https://sites.tufts.edu/emotiononthebrain/files/2014/10/Screen-shot-2014-10-24-at-11.01.01-PM-239x300.png) Hemispheric Lateralization of Amygdala Activity During Successful Recall of Emotional Memory[Credit: Canli et al. (2002)]
Hemispheric Lateralization of Amygdala Activity During Successful Recall of Emotional Memory[Credit: Canli et al. (2002)]It may be that sex differences in lateralized amygdala activity and the other regions suggested by Cahill et al. (2004) indicate corresponding sex differences in the processing of emotionally salient information. Significant evidence points to a bias of the right hemisphere (favored in male brains in emotional memory) in processing global information, such as general stimulus characteristics, and how the stimulus fits into the greater environment around it. Conversely, research points to a bias of the left hemisphere (favored in female brains in emotional memory) in the processing of local, fine-detailed aspects of the stimulus (Cahill et al, 2004).
While neither one of these strategies is inherently better than the other and the effectiveness of each is likely dependent on context and what is trying to be remembered, it suggests that women may hone in on the more detail-heavy aspects of emotional stimuli, which in turn somehow gives them an advantage in emotional memory. Other possible explanations have been proposed as well. Canli et al. (2004) suggested that women showed better memory than men for emotional events not necessarily because of different neural pathways employed, but because they had more total brain regions activated by subjective emotional experiences and by the encoding of these experiences into memory. Simply put—more activation in more places may simply indicate more encoding.
The clear sex differences in lateralization of amygdala activity are likely related to the findings discussed here that suggest women have better memory for emotional events than men, although the nature of this relationship is not completely understood. More research should explore and attempt to identify possible differences in encoding and retrieval strategies of emotionally salient information between men and women. Doing so may shed more light on the reasons for these sex differences in hemispheric lateralization in the amygdala and other brain regions and why women tend to have better memory for emotionally salient events.
References:
Cahill, L., Uncapher, M., Kilpatrick, L., Alkire, M., Turner, J. (2004). Sex-related hemispheric lateralization of amygdala function in emotionally influenced memory: an fMRI investigation. Learning & Memory, 11, 261-266.
Canli, T., Desmond, J., Zhao, Z., Gabrieli, J. (2002). Sex differences in the neural basis of emotional memories, Proceedings of the National Academy of the Sciences, 99, 10789- 10794.
Fujita, F., Diener, E., Sandvik, E. (1991). Gender differences in negative affect and well-being: the case for emotional intensity, Journal of Personality and Social Psychology, 61, 427-434.
Mackiewicz, K., Sarinopoulos, I., Cleven, K., Nitschke, J. (2006). The effect of anticipation and the specificity of sex differences for amygdala and hippocampus function in emotional memory. Proceedings of the National Academy of the Sciences, 103, 14200-14205.
Seidlitz, L. Diener, E. (1998). Sex differences in the recall of affective experiences. Journal of Personality and Social Psychology, 74, 262-271.
-

Irina Yakubovskaya wrote a new post on the site Emotion, Brain, & Behavior Laboratory 10 years, 8 months ago
 In this blog post, I will be providing neuro-scientific explanations of the processes of theatre performances that involve human actors. When performing, actors undergo an immense stress. It is almost as if in a […]
In this blog post, I will be providing neuro-scientific explanations of the processes of theatre performances that involve human actors. When performing, actors undergo an immense stress. It is almost as if in a […] -
Giovanna Castro wrote a new post on the site Emotion, Brain, & Behavior Laboratory 10 years, 8 months ago
While in most animals, sexual behavior is a straightforward path for procreation, sex in humans takes various shapes, forms, and colors. It can be by yourself (masturbation) or with many people (orgies), it can be […]
- Load More



![Hemispheric Lateralization of Amygdala Activity During Successful Recall of Emotional Memory [Credit: Canli et al. (2002)]](https://sites.tufts.edu/emotiononthebrain/files/2014/10/Screen-shot-2014-10-24-at-11.01.01-PM.png)