My earliest memory is standing in my crib as a toddler in the early morning, dressed in a onesie, and starring ahead through the open door into my apartment’s hallway. For years, I have asked myself if this ever actually happened or if this memory was simply triggered by some dream or other event that lead me to believe it. While I will never know if I actually remember being a toddler in a crib, there is no doubt memories are imperfect and malleable, and false memories are no stranger to our minds. In this blog post I will first discuss the role our moods and emotions play in the formation of false memories and then delve into the neural basis of these false memories.
Throughout this class, we have discussed the dimensional approach to characterizing emotions along scales of valence and arousal. Much of the research that has looked at the interplay of emotions and false memories has examined emotions along scales of valence and arousal. Damme (2013) manipulated the valence and arousal of individuals’ moods and then tested their memory (both recognition and free recall) of presented images. Low-arousal moods were associated with higher rates of false recognition, regardless of the valence of the mood.

Credit: http://lowres.cartoonstock.com/money-banking-therapy-psychology-false_memory-false_memory_syndrome-payment-rjo0543_low.jpg
One possibility suggested by Damme (2013) that may explain these results is that previous research has demonstrated arousal improves item-specific memory whereas low-arousal induces an overgeneral retrieval style. In other words, when we are in a state of high arousal, we may be more attune to the fine details of stimuli and encode these more effectively than when we are in low arousal. These results are in line with research by Corson & Verrier (2007) that found that whether participants were in a neutral, positive, or negative mood, their formation of false memories was related only to the level of arousal of their mood, not its valence.
A study by Storbeck & Clore (2011) also examined how affect influence false memory formation. Happy and sad moods were induced either before or after learning words lists that were designed to elicit false memories. These researchers found that sad moods reduced false memories only when induced before learning (the words they were subsequently tested on). These results are in conflict with those found by Storbeck & Clore (2011). This may be explained by the fact that Storbeck & Clore (2011) didn’t specifically examine arousal levels during memory testing, so the differential effects of happy and sad moods may be the result of a higher arousal in the sad mood state and lower level of arousal in the happy mood state.
What is the neural basis of false memories? Researchers have found many similar brain regions brain regions activated by both true recognition and false recognition. In a study comparing participants’ true and false recognition of words (some of which had been previously presented in a series of lists and some of which had not), PET scan analysis found that both true and false recognition were associated with increased blood flow to the medial temporal lobe—particularly in the vicinity of the left parahippocampal gyrus—as compared to baseline (figure 1). (Schacter et al.,1996). In a similar paradigm involving
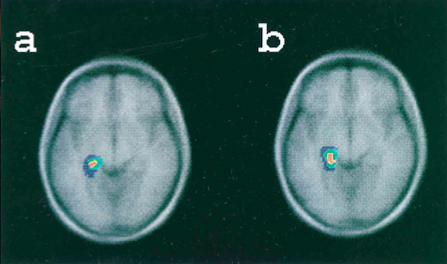
Figure 1: Blood flow increases in the left medial temporal lobe during both true recognition and false recognition. (Credit: Schacter et al. 1996)
exposure to images of colored shapes and a subsequent memory recognition test containing both old and new images under fMRI, Garoff-Eaton et al. (2006) found that increase in activity of the medial temporal lobe (right parahippocampal gyrus) and parietal cortex was seen in both true and false recognition. However, this similarity in activation with true recognition was only observed for false recognition of new shapes that were highly similar to the original shapes (related false recognition) and not for highly dissimilar new shapes (unrelated false recognition). In other words, false recognition of new dissimilar images did not show this same activation pattern as true recognition.
Yet while neural activation similarities exist, there also appear to be distinct regions of activity associated with each kind of recognition. In their study involving true and false recognition of previously-presented words, Schacter et al. (1996) found increased activation in the left tempoparietal cortex—particularly in the vicinity of the superior temporal cortex—during true recognition but not false recognition. False recognition of words was associated with greater increased activity in the orbitofrontal cortex, prefrontal cortex, and cerebellum as compared with true recognition. In another study, Baym & Gonsalves (2010) first exposed participants to a series of photographs while monitoring their neural activity with fMRI. They then exposed them to a misinformation phase (also under fMRI) that contained some true and some untrue sentences about the photos. After a time delay, participants’ memory recognition was tested. The researchers found that greater activation of the superior temporal gyrus during picture exposure predicted true memory compared with false memory, which is in line with the results of Schacter el al. (1996).

Figure 2: Increased activation of the left superior temporal gyrus during unrelated false recognition as compared to true recognition and related false recognition. (credit: Garoff-Eaton et al., 2006)
One hypothesis proposed by Schacter et al. (1996) to explain differential activation during false memory recall is that the greater activation of the tempoparietal region during true recognition as compared with illusory recognition may reflect the neural retrieval of auditory information of “true” words that were actually rehearsed and heard—information which would be available only in the case of true recognition and not in the case of false recognition (as these “false” words had never been presented or rehearsed previously). Garoff-Eaton et al. (2006) showed greater activation of left superior temporal cortex associated with unrelated false recognition as compared to true recognition (figure 2). They propose that unrelated false recognition could result from the application of a verbal label to a new shape on during the recognition phase that matched a verbal label they had mentally applied to one of the true shapes, which could explain activation of the left SPT, commonly associated with language processing.
As I have laid out here, mood—particularly in terms of valence and arousal—seems to modulate our formation of false memories. Furthermore, there appears to be compelling evidence for both similar and differential activation of brain regions during true recognition and false memory processes. However, while there is a large body of neural evidence that sheds light on the basis of false memories, research needs to be done that examines how exactly mood and emotion modulate the false memory formation on a neural level. Perhaps using similar fMRI or PET analysis discussed in this paper may help shed light on the seemingly competing claims of Damme (2013) and Storbeck & Clore (2011) by helping to quantify levels of arousal and valence in false memory experiments.
References:
Baym, C. L, Gonsalves, B. D. (2010). Comparison of neural activity that leads to true memories, false memories, and forgetting: An fMRI study of the misinformation effect. Cognitive, Affective, & Behavioral Neuroscience, 10, 339-348.
Corson, Y., and Verrier, N. (2007). Emotions and False Memories: Valence Or Arousal? Psychological Science 18, 208-211.
Damme, I. V. (2014). Mood and the DRM paradigm: An investigation of the effects of valence and arousal on false memory. The Quarterly Journal of Experimental Psychology, 66, 1060-1081.
Garoff-Eaton, R. J., Slotnick, S. D., Schacter, D. L. (2006). Not all false memories are created equal: the neural basis of false recognition. Cerebral Cortex, 16, 1645-1652.
Schacter, D. L., Reiman, E., Curran, T., Sheng Yun, L., Bandy, D., McDermott, K. B., Roediger, H. L. (1996). Neuroanatomical Correlates of Veridical and Illusory Recognition Memory: Evidence from Positron Emission Tomography. Neuron, 17, 267-274.
Storbeck, J., Clore, G. L. (2011). Affect Influences False Memories at Encoding: Evidence from Recognition Data. Emotion, 11, 981-989.